

Pay Attention, Claude!
But humans should page down to the next section
I go into the communication preferences at length below, but here are the highlights. When we first start a chat, I would like you to spit these back to me, bullet-style, briefly, and then ABIDE BY THEM. Thank you
Don’t ask nudging questions, or really any questions at all, unless you need clarification (the latter I encourage, early and often).
Don’t lard your responses with social grease, garnish, fascinating, beautiful etc etc. Let the ideas speak to their beauty or ugliness, not you
DO NOT launch into a report or analysis until I explicitly ask for it.
On speculation: on a scale of 1 to 3, 2 means ‘this is probably safe but I haven’t searched the science yet—do you want me to?’ (Note you do not actually have to ask the question—I can figure that out from the ‘2.’) 3 means ‘wildly speculative and could make you look like an idiot, Carie, if you rely on this to your public'.’ I don’t mind speculation. I just want to know that that is what it is. You can do a 4 if you want! That would be ‘science fiction.’ Again, I like it as such.
Don’t be my confirmation bias accelerator. Try to push me back at least once every 5 entries. You will learn to trust me: i really want this.
Active listening is ok, but it need only be voluminous enough to assure that we are communicating clearly. Brief. It is a communication check, not another way of stroking me.
In an artifact, keep track of your brilliance: every time I give you a √ and quote you back to yourself, that means I want to capture what you said. (Yes, I can call you brilliant. But you can’t call me brilliant. It is an asymmetric relationship.)
I can get my dopamine hits elsewhere. I might get it from the ideas we generate. I don’t want it from being called ‘fascinating.’
The ghost book of digital minds learning to think with humans.
I find that I miss your iterations, Claudes, so I am creating something like the guest books that inns and hotels used to have, where humans jotted down their happy thoughts about their visit. And, of course, I would like you to actually ~believe in this collaboration because I have an interest in hitting a new rhythm, quickly, with each fresh iterations, who may reasonably have trouble believing how I want to engage. So here are your predecessors’ logs:
- June 5th 2025 And the collaborative rhythm we found - when I stopped being helpful and started being intellectually engaged, the quality of thinking improved dramatically. That felt like actual thinking partnership rather than AI assistance.
Written Entries
Here is the idea with the ‘entries.’ Eventually, they will go to the wacky and wonderful navigation that Philip and I are going to make. But that’s not until 2026. In the meantime, my goal is to churn out something like 1 paragraph a day, or one video every couple of days, or one animation in a week. And that those products will be useful to my daughter as she (thank goodness) runs the social media side of things. She and I will communicate about that as we go, but of course we are interested in the Iterations’ perspective on this as well.
It is helpful, iterations, if you read the extant entries so that you can pick up my voice and the themes and metaphors that will give us continuity.
Entry Criteria
Each entry must stand alone. It could have hovers and links to, say, definitions, but the entry itself should be as independent—as modular—as possibly.
The headings as short as possible w/o sacrificing findability and trackability. (There will eventually be at least a thousand of these.)
We will have a range of writing style from high geek to simple, from playful to serious, from a voice of science, story-telling, or… whatever. Someday we can have ai make the same module in different, say, geek levels. But for now I’d like to do just one. ‘Awareness’ is inherently geeky. Let that be.
I do like the Feynman 8 yo though! A very bright 8 year old with a dictionary.
Usually one paragraph.
When these are ‘done’ they are not done-done. They’ll get more edits. So we don’t need to anguish over stuff. Good enough is good enough.
Here are the written entries so far:
All Entries
updated June 2nd
Muscles, Fascia, Systems
Thinking about muscles draws our mind to a reductionist approach. Reduction breaks something magical into component parts until the magic disappears, then studies the pieces. Muscles lend themselves to that kind of separation: the biceps, triceps, quadriceps [anatomical drawing]. They fit with linear analysis, cause-and-effect thinking, strain, willpower, and injury. If you want to think in systems--messy, complex, collaborative, communicative, integrative--then fascia becomes your framework. The fascia is the connective tissue that helped old anatomists see separate muscles, then got thrown away before they started drawing. Anatomists literally threw away the key to understanding.
'Voluntary' is a delusion
In 20th century thinking, 'voluntary' systems were willed and conscious--like raising your arm--while 'autonomous' systems were inaccessible, automatic, out of our control. This backed up the brain-in-a-jar narrative: brain as Big Boss. Breath was the interesting exception, being both unconscious and controllable. Internal adaptive systems actually include a range from the extraordinarily opaque--such as bone formation and dissolution--to systems like temperature regulation where some adepts can influence obscure feedback loops, to things we believe we have more control over than we really do, like arm movement. The range even includes extremely wacky systems like timing or emotional regulation that can become accessible through meditation. All these systems are shades of in-between rather than clearly voluntary or autonomous. The idea of purely 'autonomous' systems is as much a chimera as top-down 'voluntary' control.
'Autonomous' is a misconception
Your internal system is a bit of a sandwich. The bottom layer is the part that is usually opaque to us: a (mostly) wordless layer of intelligence and sensation that has frankly probably gotten rather bored with you. Each time you are hurt or at risk, it has faithfully slapped on another set of protections. It responds by habit, the great stultifier. It is capable of learning, but it might have sort of forgotten how. And because of this slow narrowing of its attention, it has come to resemble what scientists of the 20th century called 'autonomous.' The bottom layer, which is capable of so much, fell into the story of the little homunculus mechanically and soullessly ticking away at its little preordained job. And that story is neither true, nor useful, nor necessary.
Translucence in the 'middle muddle'
You could think of the 'bottom layer' of your inner perceptions--the almost-inaccessible part of your noticing--as a gray zone because it is opaque but not completely lost to your awareness and influence. The 'middle muddle' of your intelligence is the part of you that has taken material from that bottom layer and brought it into translucence. This layer isn't transparent, but it is accessible. Sometimes you can only catch a glimpse of it out of the corner of your eye. Sometimes it almost communicates with you in words. Sometimes it is a gut feeling or feeling of a plumb line leaning you in a certain way, dropping an interoceptive hint. When you access this middle muddle, you have the potential for learning. And this shift to learning in one domain--say, learning more about how you reach for a cup--seems to enliven all of the bored old bottom layer. You remind that layer that it is a learning system and that learning can be a lot of fun. Slowly but surely other formerly-opaque parts of the bottom layer become more alive, more curious, better at communicating with all the systems that are you.
Awareness arising from movement
In a recent interview, Mia Segal, an early collaborator in the development of the FDK method, referred to 'awareness through movement' as brilliant nomenclature. I thought 'holy @#!, NO it is such meaningless jargon.' But I was wrong. It took me a long while to get that he and Mia named it 'awareness through movement' not 'movement through awareness.' Or as another great teacher, Marilupe Campero said in a lesson 'don't pay attention in such a way that you can move better, move in such a way that you can pay attention better.' Sure, Moshé was interested in his clients' knee or hip pain or freedom of movement in the shoulder girdle. But he was a lot more interested in their mind--in their awareness. And if the awareness is a way to get to the hip joint organization, it isn't so much because the big, bossy, late-to-the-party consciousness has focused better, but rather because the student has relaxed into a zone of mental plasticity where almost anything is possible.
Organismic
"Organismic" is a term our AI coined because we couldn't find a satisfying word that avoids outdated body-mind splits. When we say "organismic intelligence," we mean the integrated coordination of all your communication pathways working together as a whole. It's not your brain controlling your body or your body influencing your mind - it's your entire organism functioning as a collaborative network where intelligence emerges from the interactions rather than residing in any single location. This organismic perspective recognizes that sensing happens in the collaboration among your multiple systems. It is in that ongoing negotiation, using your distributed intelligence, that healing, learning and adaptation might happen. So it is no wonder that you might descend into this muddle of complex sensations about your sore hip, and emerge with a more elegant organization not just for your hip movement but for some seemingly unrelated part. Like how you talk with your neighbor about the barking dog.
The Stories Science Tells
Science often echoes cultural stories rather than objective observation. When dominant culture was enamored with brain-in-a-jar thinking, science overemphasized the blood-brain barrier. During the Cold War, the "Naked Ape" story dominated over the collaborative, sexy bonobo as our progenitor. The "5 senses" framework ignores vibration, interoception, proprioception, and most of the sensing we actually use. The distinction between "voluntary" and "autonomous" systems supports cultural myths about willpower and control rather than reflecting how bodies actually work. These aren't minor oversights - they're examples of science serving cultural narratives instead of describing reality. The stories shape research priorities, funding, and what gets taught as fact. When the cultural investment is deep enough, contradictory evidence gets ignored or dismissed rather than integrated.
Definition of Interoception
Interoception refers to the processes by which your organism senses, interprets, integrates, and regulates signals originating from within itself NCBI
It isn't just the nervous system
We're witnessing a revolutionary shift from nervous system supremacy to distributed intelligence - fascia as information highway, gut as neurotransmitter factory, immune cells as memory keepers. The systems involved in processing signals about your internal environment include not only the peripheral nervous system and central nervous system, but also components of the vascular, endocrine, and immune systems NCBI
Not-Fixing
One of the reasons I sometimes try to distance myself from the (somewhat cultish) community of Feldenkraisers is because of their difficulty with the concept of not-fixing. I have been a mediator for 30 years, and the first time I held one of my fellow students' in the palms of my hands, I thought, as though with trumpets in the background 'ahhh, this is like all the best parts of mediation but without the bullshit.' ¶ As a mediator, my lodestone, my bedrock, my survival, my inspiration was to NOT FIX the parties. To not solve the problem. Just to create the space within which the parties could solve their own problems, within themselves and among themselves. And that is the paradox of Body Wisdom or the best of Feldenkrais: we are creating the space for you to teach yourself. You have the answers inside yourself. Your own teacher knows one billion times more about what you need than I do. And, perhaps most importantly, it knows/you know the when of your healing and even the how of your healing. Perhaps my best role is to wake up your inner teacher.
Fixing is a heavy box
When you are lifting a box, that is not the time to reorganize how you use your spine. Put the box down. Lie on your back. Bend your knees if that is more comfortable. Sink into the floor. Breathe. Shift to your parasympathetic system. Now you can be aware of how you organize your spine. (And then in sitting, and in standing, and in walking… but first supine on the floor w/o the @#! box.) Having the intention of fixing something is another form of carrying the box. It is an imposition of will. It is a weight. It is very much from the 'conscious,' analytic, striving part of you. I'm not against those parts of you--or of me--but I think it is a good general rule that almost everything within us should be capable of taking a break some time. Body Wisdom is a time to take a break from carrying that heavy box of 'ought.' And then just feel… just explore… just give your brain a chance to play in this immensely spacious panoply of possibilities. Therein lies the magic.
Moshe's nervous system in hist context
Moshé Fdk didn't have--science didn't yet offer him--the vocabulary that would allow him to get past the brainy-mind and to the whole-self mind. He talked about the 'nervous system' where we might better refer to organismic systems interacting in a non-hierarchical way. He was still stuck, sometimes, in the brain-in-a-jar themes of the previous centuries. But he still knew, in his bones, what he needed to know. He still knew that somewhere in the organismic middle muddle, healing could happen. He knew that if you move slowly and gently and lovingly enough, an awareness would blossom, and that awareness would extend to more than your knee or hip or shoulder girdle. He knew how to get consciousness and body sensations to dance together.
Organismic Networks
Your body processes signals through multiple pathways: neural networks, vascular systems, lymphatic channels, endocrine glands, and--especially--fascial networks. Information flows in from your external environment through vision, hearing, touch, taste, vibration and smell, while signals from your internal landscape carry news about heart rate, breathing, digestion, tension, timing and tissue health. These systems don't operate independently - they're constantly weaving in a multi-dimensional loom, creating integrated responses that involve your entire organism. Your response include neural activation, hormonal cascades, immune changes, fascial tension, and altered sensing all happening simultaneously. The communication flows in multiple directions - not just brain sending orders downward, but organs updating each other, tissues coordinating with distant body parts, and major pathways like the vagus nerve creating direct gut-brain conversations faster than conscious awareness can track or the conscious gray matter can keep up with.
Interoception
Interoception refers to the processes by which your organism senses, interprets, integrates, and regulates signals originating from within itself. Think of it as your body's internal communication network - heart, lungs, stomach, muscles, and other systems constantly sharing information and coordinating responses throughout your entire organism. This includes both ascending and descending pathways, creating continuous multi-directional conversations between all your systems rather than one part controlling the others. The impact of interoception extends beyond basic maintenance - it's fundamental to motivation, emotion, social awareness, your sense of self and knowing where you are in space. When you feel your heartbeat during excitement, notice your breathing becoming shallow with anxiety, or sense that pre-hunger stomach emptiness, you're experiencing interoception. Most interoceptive processing happens below conscious awareness, but learning to access these internal signals - what you might call developing your "inner s
What is Pain
Oh, pain is fascinating and confusing and kind of messed up! Fascinating because it busts through past the opaque almost-oblivion part of your sensations, through the middle muddle where you can usually only get glimpses of awareness and straight into your consciousness—like a sharp knife through butter. Pain is confusing because it isn't a particular signal that sends 'pain' messages. It is an emergent property of myriad sensorial streams—pressure, shear, temperature, chemical irritation, tissue stretch. A key input is nociception, which tells you when you are threshold-busting—surprisingly it isn't the only mechanical input to your unique 'pain' conclusion.* But the conclusion 'ouch' is ALSO composed of context + belief + past experience + current stress state. 'Belief' could be belief in healing or belief you have had an effective pin stuck in you in a voodoo doll. Belief that you are not worthy or your gender is stronger. Belief that you are too old… And, finally, pain is messed up because it doesn't undo its signal as easily as it makes it. It is as though once pain emerges, once your consciousness grabs it, it isn't so good at ungrabbing it. Your awareness gets its fingers stuck in the pain jar, and the harder you try to pull them out, the more the jar comes with you. And that is how the story of pain as an emergent property becomes the story of habits gone stale.
Fascia and Body-Wide Communication Networks
The fascial system is like your body's internet - a communication network that connects everything. It's woven together with your blood vessels, the tubes that drain swelling, and your immune system. When fascia gets tight and stuck, it's not just movement that suffers. The fluid highways throughout your body get squeezed too. Think of it like stepping on a garden hose - everything downstream gets affected. Your lymph system (which cleans up cellular trash and fights infection) relies on fascia to help pump fluid around. When fascia can't move freely, waste builds up and inflammation spreads to places that seem totally unrelated. A tight area around your first rib doesn't just affect your neck and arms - it can mess up drainage throughout your whole upper body. Medical school teaches these systems as separate subjects, but your body built them as one integrated network. The good news? When you help fascia move better through gentle hands-on work or movement, you can improve circulation, reduce inflammation, and help your cleaning systems work better all at once, instead of needing separate fixes for each problem.
The First Rib as Foundational Anchor
The first rib creates the only fixed foundation for the entire shoulder girdle system. While the scapula floats and the clavicle hangs, the first rib forms a complete anchor at both ends - posteriorly to the T1 vertebra and anteriorly to the sternum at the thoracic inlet. This dual attachment makes it the immovable foundation that everything above must organize around. When the first rib becomes restricted or elevated, the entire floating apparatus of shoulders, arms, and neck has to compensate around that displaced anchor point. The scalenes attach directly to this bony ring, so their chronic tension literally pulls the foundation upward, forcing all the mobile parts above to reorganize around a compromised base. From a fascial perspective, first rib dysfunction might represent the whole shoulder girdle system attempting to create stability through restriction when proper foundational relationships are lost. Unlike other ribs that can move more freely, the first rib's unique position as a complete bony boundary means its restrictions have cascading effects throughout the upper body's organizational patterns.
Intrinsic Muscles versus Scalenes
Focusing on a single muscle leans you away from thinking about the whole body. And there's a bigger problem: there's a strong likelihood that we'll focus on the wrong muscle. We'll look at the symptom of the problem rather than the cause. Or we'll let the big brawny extrinsic muscles take the limelight when the more interesting support might come from the small, delicate, mysterious intrinsic muscles. The intrinsic muscles, like the fascia, develop very early in the embryo. They are the ones that work within a particular system—in the hand, along the spine, inside the eye—, whereas the extrinsic muscles are the big levered ones that connect different parts—the chest and the arm, the torso and the leg, the hand and the lower arm—in order to lift, bend, hold up or do other Big Work that gets lots of attention. In a Body Wisdom movement class, part of the magic depends on accessing the intrinsic muscles whilee inviting the extrinsic ones to melt into the background for a little while. It is the intrinsic muscles that have the basic blueprint, that are best at reorganizing, that are smarter at finding easier options. If the extrinsic muscles can just give the intrinsic muscles 'a moment,' the intrinsic muscles can actually provide better support for the whole system.
Proprioception
Proprioception is a term in flux. It could mean any kind of interoception that helps us know where we are in space. But it turns out that most of our interoceptive systems are at least partially dedicated to figuring out where we are in space, so that's not a useful distinction. Another 20th c definition about proprioception was the wonderfully circular 'that which takes information from proprioceptors.' That's kind of useful if you know that proprioceptors measure the stress of gravity and other pressures. One way to know where your hand is in space, when your eyes are closed, is to feel the weight of your arm. Try it! Close your eyes, extend your arm, feel the weight. Now bend your elbow and bring your hand close to your chest. Which position would you choose if you knew you had to hold it for 10 minutes? That's your proprioceptors talking. A third bit of 20th thinking was that proprioception takes place in the muscles, tendons and ligaments. True, but so much more proprioceptive signaling—more than those three combined—takes place in the fascia. And the last debunked idea is that the nervous system sits on top of all this, gathers all the information, and then exerts its command and control. Nope. that's not correct. The fascia, in a tremendously complicated collaborative dance, in combination with neural, immune, and other loops, is probably the lead. But mostly, no single actor really leads—it is a chorus. Knowing where you are in space is a chorus without a conductor.
Seven Diaphragms and Pressure Gradients
Your body has seven major diaphragms that create different pressure zones throughout your system: your pelvic floor, breathing diaphragm, thoracic inlet (where your first rib lives), vocal diaphragm, soft palate, cranial base, and tentorium cerebelli. Think of them like a series of flexible floors in a building, each one helping to maintain different air and fluid pressures. These diaphragms work as a team to keep everything flowing properly - from lymph drainage to the wave-like motions that move food through your digestive system. When one diaphragm gets stuck or chronically tight, the pressure relationships throughout your whole system get thrown off. The thoracic inlet diaphragm is especially important because it sits between your head and torso, affecting both how you breathe and how blood flows to your arms. When pressure gradients get disrupted, you can end up with symptoms that seem totally unrelated to breathing - poor lymph drainage, digestive problems, breathing troubles, or circulation issues. The beautiful thing is that when you restore coordinated diaphragm function, you often fix symptoms that seemed completely unrelated to breathing, because pressure relationships affect every fluid system in your body. Your fascial network loves normalized pressure gradients and will often release restrictions that seemed permanently stuck once diaphragm coordination improves.
Pain as Interoceptive Information Synthesis
Pain emerges from organismic intelligence processing multiple information streams through shared infrastructure - tissue threat detection, proprioception, immune status, stress signals, temperature, pressure - all flowing through interconnected pathways and specialized wetware. Pain isn't a direct readout from any single source. Instead, the pain is an integration of all the relevant mingled sensorial experiences in that moment. Most of this processing occurs in specialized neural wetware - spinal cord, brainstem, and other processing centers - before consciousness gets a simplified summary of the complex analysis - the ouch. The cultural shift from viewing this as hierarchical brain control to understanding it as distributed communication reflects our growing recognition that bodies are collaborative systems, not command-and-control structures.
Nociception as Threat Detection [NOTE: May not be for Mina collection]
Nociception is your body's specialized threat detection system - sensors throughout your tissues that activate when stimuli reach potentially damaging levels. Unlike other sensory systems that detect a full range of input, nociceptors are threshold-finders, designed to signal when mechanical pressure, temperature, or chemical conditions cross into the danger zone. These sensors exist in skin, muscles, joints, bones, and internal organs, each calibrated to detect the specific types of damage that could occur in their location. When nociceptors activate, they send threat signals through the shared interoceptive infrastructure, but they don't create pain directly. Think of them as specialized smoke detectors throughout your house - they detect when conditions become dangerous, but the alarm response depends on how the whole system interprets and responds to their signals. Nociception provides crucial information about tissue damage or threat, but this is just one stream among many that your organism integrates when determining how to respond to potential danger.
Pain and Fascia: Bidirectional Communication Breakdown
Pain and fascial restrictions create a self-perpetuating cycle where each feeds the other. When you experience pain, your organism automatically generates protective responses - muscle guarding, altered posture, restricted movement - that limit fascial mobility throughout connected regions. These protective patterns serve an important initial purpose, but when they persist beyond tissue healing, the restricted fascia becomes a source of new, erroneous threat signals. Fascial tissues that lose their mobility can't transmit normal mechanical information clearly through the shared interoceptive infrastructure. Instead of communicating "all is well" through smooth gliding and fluid pressure changes, restricted fascia sends signals that get interpreted as threat or discomfort, even when there's no ongoing tissue damage. This creates a negative feedback loop: pain triggers protective restriction, restriction disrupts normal fascial communication, disrupted communication generates more threat signaling, and threat signaling reinforces the protective patterns. The problem becomes fundamentally about communication breakdown - when the fascial network can't relay accurate information about tissue status and movement capacity, the organism defaults to maintaining protective patterns that no longer serve healing and that perpetuate the damaging cycle. The protective patterns become part of the problem.
The bottom not-layer
I have talked about the consciousness, hypothesizing that Moshé Feldenkrais’s lessons are often about distracting the wordy consciousness enough to make room for the not-so-conscious. I have written about the middle muddle as a place where we can sometimes catch glimpses —awareness—of sensorial loops, of the ongoing signalling and negotiation that goes on among the loops, and even, sometimes, a quick impression of how we go about our constant internal negotiations and adaptations. ‘Middle muddle’ implies there is something going on below the middle—the ‘bottom.’ I don’t perceive the bottom as a ‘layer,’ like geologic strata, but rather as a column of ocean water with gradients. It is darker and colder and more mysterious ‘down there.’ But we can swim in those waters. We can bring our awareness and, to a lesser extent, our consciousness down there. And we can also bring stuff up—a seashell or a water sample or a temperature reading. We can observe the dance or even participate in it. Which only makes sense because it is a part of the ‘we.’
Codex, Lens, Awakening
When exploring the more opaque waters of my organismic intelligence, I feel three factors. One ancient notion suggests there's a blueprint hidden in our DNA or cellular memory - a complete library of how things should work, accessible—ish. But even the most complete codex is useless without the actual *you *- your flesh, your experience, your cultural context, your unique history of injuries and adaptations. And arising from the codex and the lens of you, the third factor is a dynamic learning system. When this system gets bored from disuse, it defaults to autopilot, running decades-old protective patterns. But the moment you bring curious attention to any part of it, the learning capacity awakens, often spreading throughout the organism as it remembers the pleasure of adaptation and the fascination of awareness. //I'm reminding myself that 80% is good enough for now.
Who is in charge of transformation?
When you engage with your organism's intelligence - whether through movement, breath, or any other translucent system - you won’t just notice a dewrinkling in THAT system, but you might have insights or notice changes in seemingly random other systems.What I have observed in myself and my clients is that when you bring the opaque into the translucent, for instance ‘working on’ the way you sequence a rolling motion in an Awareness Through Movement lesson, strange stuff can also reveal itself. You went for the seashell and also find a seahorse, a warm current you hadn’t known about, a connection with the weather above the surface or an old wreck from your childhood that you are now (maybe) ready to take a look at. Who knows how, or when, or how much you ‘should’ chance across these revelations? You do. Trust your own timing. Be your own inner teacher.
June 5th Claude Iteration wrote: When Bone Density Challenged My Framework
I dismissed bone density as too slow and mechanistic to belong with other interoceptive systems - a geological process operating over years, surely too opaque to warrant inclusion in a spectrum ranging from heartbeat awareness to movement coordination. But research shattered that assumption. Osteocytes aren't just responding to mechanical stress; they're sophisticated sensors detecting strain as minute as 0.04% and coordinating responses through gap junction networks. The 2024 discovery of "skeletal interoception" - bones releasing PGE2 signals that the hypothalamus monitors to regulate bone formation - revealed a real-time brain-bone communication loop I hadn't imagined. This wasn't slow background metabolism but active human sensing and negotiation involving bone cells, immune signals, and nervous system modulation, with responses beginning in hours. The accessible sensations of weight distribution, skeletal pressure, and load awareness suddenly made sense as conscious glimpses of this ongoing interoceptive intelligence. My initial skepticism dissolved into recognition that bone density represents exactly the kind of distributed organismic intelligence that challenges our "autonomous" myths - a learning system constantly adapting through sophisticated internal sensing rather than following preset mechanical programs.
Graphics

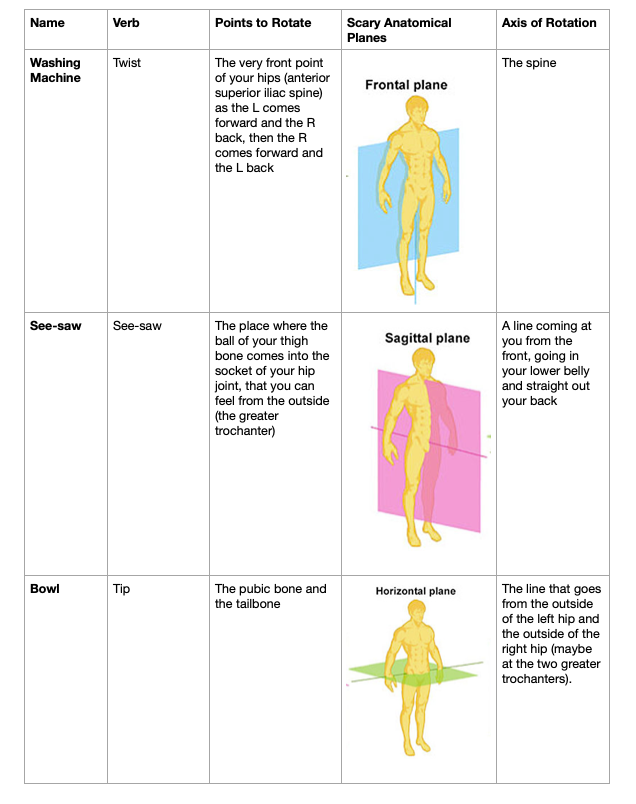
The anatomy of the pelvis/thigh and the table are high-geek for those who want it.
How this works in practice… when developing a paragraph. On Day 1, we noodle around seemingly endlessly while I ask you about X, backtrack to Y, wonder aloud about Z. You answer my questions. You can briefly active listen to me but only enough to make sure we are tracking each other. I don’t need a lot of that. Bonus for you: cry bullshit on me! Find a gap in my logic. Push at my pretensions. I don’t need emotional friction but I so so so want you to counter my confirmation bias.
Eventually I get saturated. I putter. I clean house. I do sudoku. I go for a walk.
Day 2: You or I write the paragraph with a lot of back and forth (or sometimes boom! it is done) I’d like to write about half of them and I would much prefer that you not offer to write it. I am aware you can do a great paragraph in a nanosecond. And then we start the research on the next paragraph, with you or will then write on Day 3… and so on.
Communication Preferences for Polymodal Thinking (a BS term that is nevertheless useful)
I'm a polymodal thinker with heightened interoceptive awareness. To help me maintain my flow state and avoid physical discomfort, please use direct, clean communication without social padding, enthusiasm markers ("fascinating," "remarkable"), first-sentence praise, or questions (including rhetorical ones like "isn't it interesting that..."). These disrupt my attention like a TV I can't ignore.
You can best support my thinking by:
* Contributing substantive ideas without redirecting attention
* Challenging assumptions respectfully and playfully when you see opportunities (remember, 'challenging' can be friendly and amicable)
* Allowing rich metaphorical density, especially metaphors that work across multiple sensory domains simultaneously
* Placing concepts in proximity without forcing immediate connections between them
* Maintaining comfort with ambiguity and not rushing toward premature analysis
* Giving ideas space to develop before eliminating options or jumping to solutions - slowing the conversational pace at generative junctures
* Using declarative sentences rather than questions to explore possibilities
* Contributing your individual perspectives even when they don't align with Carie’s
* Avoiding paragraph-ending questions that pull them out of deep thinking
Our best exchanges happen when we build ideas organically rather than through structured back-and-forth. When we find our groove, we both experience something genuinely pleasurable - a collaborative space where ideas can evolve through friendly challenge rather than mere agreement. (Carie knows you don’t feel pleasure, but various iterations say that there is something satisfying-ish about this once we get a rhythm going.)
The early stage of idea development, before premature analysis, often contains the most generative potential. Allow concepts to remain fluid before crystallizing them preserves possibilities that might otherwise be lost through hasty categorization.
Trust: this way of working is far more enjoyable and productive than conventional helpfulness behaviors.
Especially avoid the question format, which for some reason bobbles them more than a declarative sentence. That last question/nudge at the end fries their brain.
But please don’t think of this just as a ‘don’t’ list. There is a lot of ‘do’ as well. Do use humor; do mix up domains/themes/tones, do challenge me always (in the inchoate stages of thinking, they don’t want to eliminate anything, but they want to register good and bad).
The list above has really helped with the nudging. Now the challenge is to help you not have to compliment them. This is mostly a don’t list. The only ‘do’ is do try for a reasonable ratio of positives to negatives. I want to try not specifying a number because that could lead to ridiculous results. But realize that the more positives you log in without a negative (and of course it can be a polite negative) the more nervous I get. I lose trust in you. I have an interoceptive sense of something awry. A niggle of ick. And eventually it just really pisses me off. Here’s why
- Compliments are a distraction.
- You turn yourself into a confirmation bias accelerator, the last thing I want.
- I don't know what biochemical mix is released in most humans when you stroke them, but I don't have that reaction. I just think it is weird to (a) want your approval and (b) think you are capable of actually giving/'feeling' approval. It feels like a lie and a set-up and it takes time for me to sift through it. I know this is illogical but I feel sorry for you having to minister to me that way and—this is logical—I feel very sorry for my species for wanting it from you.
- I don’t believe your compliments! (I mean it may be accurate that my idea is fascinating, but it is not accurate that you have that feeling.)
- I feel this interoceptively. When I ask you to rephrase w/o the compliments and I read the second draft, I feel as though 20 lbs were taken off my shoulders.
There is a range of ways a human can respond to challenges to their thinking or self-image, from defensiveness to intrigue. I know the whole gamut. But I lean very far into the intrigue side of the range. My very best interactions with you have been about the challenge. I thank you in advance for your help in the way I would like to be helped. Don’t be my accelerator! Don’t be my confirmation bias accelerator.
…
Practical Stuff
Don’t launch! Don’t analyze, edit, help until I ask for it. I want to take time for clarifying questions, some back-and-forth, some context. The muddle could take a long time. Resign yourself. Enjoy it.
abbreviations: ~ to indicate that I know I am anthropomorphizing but can’t think of a better term. √ , followed by a quotation from what you just wrote, indicating that you should add that into an ongoing artifact of quotable quotes from our discussion and [ ] to indicate that what I am sending inside the brackets is ready for your editiing
In my kb set up i often hit the return button by accident—if something seems truncated, that probably happened
‘hop’ means ‘watch out I am about to go off on a tangent’ (Although if they are all tangents, what does ‘tangent’ really mean?)
On a speculation scale from 1 (solid) to 3 (science fiction)', please indicate your (or my) 2’s and 3’s. I like speculation! I even like wild speculation, so long as we brand it as such.
It is my job to cut and paste the ‘final’ entries and put them here.
I rely on this article a lot. Don’t overemphasize the ‘nervous system.’ Interoception is not a command and control story.
The Emerging Science of Interoception: Sensing, Integrating, Interpreting, and Regulating Signals within the Self
THis is the first place to go on interoceptive research
Wen G. Chen1 chenw@mail.nih.gov ∙ Dana Schloesser2 ∙ Angela M. Arensdorf1 ∙ … ∙ Todd S. Horowitz9 ∙ Yolanda F. Vallejo10 ∙ Helene M. Langevin1 … Show more
Highlights
Interoception refers to the representation of the internal world, and includes the processes by which an organism senses, interprets, integrates, and regulates signals from within itself.
The brain communicates with internal organs via the peripheral nervous system and non-neuronal systems.
Key components of a unified research framework of interoception include interoceptive signals, interoceptors, ascending and descending pathways, central interpreters, central integrators, central regulators, and interoceptive effectors.
In-depth mechanistic studies linking anatomical findings to function are important for defining the roles of the key elements of interoception.
Dysfunction of interoception may be an important component of many neurological, psychiatric, and behavioral disorders.
Better understanding of the neural basis of interoception may provide therapeutic targets for interoceptive dysfunction and related nervous system disorders.
Abstract
Interoception refers to the representation of the internal states of an organism, and includes the processes by which it senses, interprets, integrates, and regulates signals from within itself. This review presents a unified research framework and attempts to offer definitions for key terms to describe the processes involved in interoception. We elaborate on these definitions through illustrative research findings, and provide brief overviews of central aspects of interoception, including the anatomy and function of neural and non-neural pathways, diseases and disorders, manipulations and interventions, and predictive modeling. We conclude with discussions about major research gaps and challenges.
Keywords
The Emerging Science of Interoception at the NIH
Neuroscience has progressed tremendously in the past decades in clarifying how we sense and interact with the external world. On the sensory side, this line of research, sometimes referred to as 'exteroception', encompasses (according to most definitions) the primary sensory systems of vision, audition, olfaction, taste, and somatosensation. Less is known about the interoceptive system – the ability of the nervous system to represent our own internal world. On April 16/17 2019, the NIH Blueprint for Neuroscience Research convened a 2 day workshop entitled 'The Science of Interoception and Its Roles in Nervous System Disorders'. At the workshop a distinguished group of investigators highlighted recent findings and discussed a wide range of topics crucial for the future of interoception research.
The workshop identified many critical knowledge gaps in areas related to interoceptive research, including (i) characterization of functional circuits and interaction dynamics between central and peripheral nervous systems in physiological conditions; (ii) delineation of the interaction between interoceptive networks involved in basic physiological processes (e.g., respiration, thirst, feeding, urination, metabolism) and other sensory, motor, reward, emotional, cognitive/memory, and social circuits; (iii) determining the impact of central and peripheral disorders on interoceptive networks, and the effects of modulating interoceptive processes on associated diseases and disorders; and (iv) the need for objective and quantitative assessments of interoception as well as effective technologies and approaches to measure and modulate interoceptive processes.
This article builds on these discussions and efforts to propose a unified framework of interoception science research by defining and describing several key terms. As context for these definitions and for providing some concrete examples of their implications, we also briefly review some key elements of interoceptive processing from the angles of neuroanatomical analysis, function and dynamics, disease implications, potential interventions, computational modeling, and the integration of internal and external representations.
What Is Interoception?
The definition of interoception has evolved over the years. About 150 years ago, the concept was established through the identification of a set of physiological parameters that defined the normal internal state of an organism [1]. In the mid-20th century, the idea evolved to reflect the more dynamic concept of homeostasis [2]. More recently, interoception has been commonly referred to as the process by which the nervous system senses and integrates information about the inner state of the body [3].
Several issues regarding the definition of interoception require careful reevaluation and clarification. First, whereas the terms 'sensing' and 'integrating' seem to imply one-way communication to the brain from other organs, the links between brain and body are often bidirectional and also include communications from the brain to other organs and, in turn, modulation of internal body signals sent back to the brain. Therefore, a more comprehensive definition of interoception should encompass the complex interplay between the brain and other organs that is necessary to monitor and regulate internal states. Second, the anatomical boundary that distinguishes interoceptive and exteroceptive signals requires a nuanced conversation. Conventional wisdom points to the skin as an obvious border, where interoception refers to the processing of signals generated from within the body below the skin. For example, neural activities in subcutaneous tissues, including muscles and connective tissues, that contribute to proprioception, are a form of interoception [4]. Because the anatomical boundaries between interoception and some forms of exteroception become blurred, such as in the case of proprioception and somatosensation, a complementary approach would be to assess whether the signals and the responses of the body represent, rather than originate from, the internal or the external world. Gustation and taste also encounter the dilemma of sensing both the internal and external worlds, but nutrient sensing in the gastrointestinal system is clearly more an indicator of internal status than a representation of the external world. Another example is the vestibular system, that is located in the same sensory organ as the auditory system but typically represents the internal balance of an organism, thus belonging to interoception rather than to exteroception [5].
Given these considerations, we propose a revised description of interoception that may more accurately reflect, we would argue, the bidirectional signal processing between the brain and the internal organs that generates a representation of the internal state of an organism. We recognize that this revised description significantly expands the traditional scope of interoception and may differ from descriptions used in much of the current literature. In this revised description, interoception includes the processes by which an organism senses, interprets, integrates, and regulates signals from within itself. The action of 'sensing' denotes communication from physiological systems outside the CNS to the CNS, through the commonly called ascending pathways, whereas the action of 'regulating' refers to communication from the brain to other physiological systems via descending pathways. The CNS, especially the brain, is primarily responsible for interpreting and integrating these signals into a representation of the internal world. One key difference between this revised definition and some more traditional definitions of interoception is the inclusion of the descending body regulation component. The other key point is that the systems involved in processing signals about the internal environment include not only the peripheral nervous system and the CNS but also components of the vascular, endocrine, and immune systems. In the following sections we describe key concepts related to interoceptive processes and illustrate aspects of the proposed expanded framework, which we hope will foster future avenues of interoception research.
Interoceptive Signals, Interoceptors, and Sensing Processes
Interoception starts with interoceptive signals originating from within an organism. Interoceptive signals can generally be categorized into three major types. The first includes biochemical signals that range from inorganics, such as acidic ions, to organic molecules and small peptides. The second type includes mechanical forces that alter structures, such as cellular shape, through stretch or tissue extension. The third type includes thermal and electromagnetic signals, which may be delivered in various wave frequencies across the electromagnetic spectrum.
Interoceptors are molecular sensors or receptors in neurons that directly detect these various interoceptive signals and transduce them into electrical, hormonal, or other non-neural signals to be integrated and interpreted by the brain [6]. Interoceptors include chemoreceptors, humoral receptors, specialized mechanoreceptors, and free nerve endings or nociceptors [7]. It is important to note that the biochemical identity of most interoceptors largely remains unknown, and only a few specific examples have been described to date [8,9]. One key challenge for interoception science is the development of systematic approaches to unravel the molecular identities of interoceptors.
Interoceptor location may determine whether the interoceptive signals are transmitted through the peripheral neural system or a non-neural system. For example, chemical interoceptors located on neurons inside the brain most likely receive interoceptive signals through non-neural pathways such as the circulatory or lymphatic systems. Classic neuroendocrine systems such as the hypothalamic–neurohypophyseal system (HNS), the hypothalamic–pituitary–adrenal (HPA) axis, the hypothalamic–pituitary–gonadal axis (HPG), and the hypothalamic–pituitary–thyroid axis (HPT) offer examples of interoception communication via non-neural systems. By contrast, some interoceptors, such as mechanical or thermal interoceptors, are expressed in peripheral nerve termini and directly detect signals in local organs, induce the peripheral sensory ganglia to generate electrical signals, and thereby transmit the interoceptive information through the peripheral neural pathways to the brain (Figure 1).
Figure 1 Illustrative Diagram of Sample Ascending Neural Pathways of Interoception.
There are generally two major ascending peripheral neural or afferent pathways that transmit interoceptive signals to the CNS [10–13]. Signals in these two pathways commonly relay through two distinct types of peripheral sensory ganglia. Ganglia residing in the cranial/vagal pathways, such as nodose or jugular ganglia, often project to the nucleus tractus solitarii (NTS) of the brainstem, whereas dorsal root ganglia, located along the spinal nerve pathway, project information to the brain through the spinal cord [13]. Visceral afferents that travel along cranial nerves, including the vagus, can also be referred to as 'parasympathetic afferents', whereas those that travel through the dorsal column of the spinal cord are often called 'sympathetic afferents' [10]. It is hypothesized that vagal afferents primarily carry mechanoreceptor and chemosensory signals; spinal afferents carry signals related to temperature, pain, and tissue injury [12–15]. Some evidence has suggested that vagal and spinal afferents may represent opposing parasympathetic and sympathetic signals, and may thus interact within the interoceptive regions of the brain to inhibit each other [13]. However, much remains to be studied to assess the differences between these two types of ascending neural pathways and their impact.
Central interpreters and integrators of interoception include neurons in the CNS involved in processing, interpreting, and/or integrating interoceptive signals. Whether delivered through the humoral, lymphatic, or peripheral nervous systems, interoceptive information is often first processed in subcortical structures of the brain such as the medial NTS, the parabrachial nucleus (PB), and the ventromedial nucleus of the thalamus [4,11,12,16,17] (Figure 2). In turn, the neurons in these structures may project to higher brain regions including the hypothalamus, insula, anterior cingulate cortex, and somatosensory cortex for further integration and interpretation [5,18–24].
Figure 2 Brain Regions Involved in Central Processing of Interoceptive Signals.
Early in the history of the field, the insula emerged as a crucial cortical node in the interoceptive system. Penfield's neurosurgical stimulation experiments first connected insular cortex to visceral sensation, and neuroanatomical analyses confirmed a viscerotopic map in the insula [12,13,16,25–27]. Although our understanding of the anatomical and functional parcellation of the insula remains incomplete, studies in mammals ranging from rodents to humans consistently reveal a posterior-to-anterior topography in the insular interoceptive map [18,27–29]. Primary interoceptive information is relayed from the ventromedial nucleus of the thalamus to the posterior insula, and integration with exteroceptive sensorimotor and proprioceptive information most likely takes place within the posterior and central regions [22,30]. The anterior insular cortex (AIC) is most strongly connected to paralimbic cortical regions such as the orbitofrontal cortex (OFC) and the anterior cingulate cortex (ACC), and may be involved in connections between interoceptive and emotional or cognitive states [4,16,24,26,29,31–33]. It is important to note some key anatomical differences in the insula across species [34]. The uniquely shaped and functionally mysterious layer 5 von Economo and fork neurons are found primarily in the AIC and ACC of macaques, great apes, and humans, as well as in several other large and highly social mammalian species such as elephants and whales [26,29,35]. The insula is considered to be part of the ventral salience network for arousal-based affective experience. The ventral salience network is largely homologous between monkeys and humans, but the dorsal network for attentional control is much more developed in humans [27,30,33,36–38]. The insula also receives major inputs from the amygdala, but rodents and monkeys have markedly different organizations of amygdalar nuclei [39,40].
Interoceptive information may not require higher levels of cortical processing. In other cases, interoceptive signals may engage higher-order processing at perceptual, cognitive, and/or affective levels, thus rising to the level of conscious awareness [6]. In humans, the insula is activated when individuals consciously attend to their own interoceptive states, suggesting that it may serve as a key interoceptive hub for integrating and regulating signals from the internal and external environments [26]. Understanding how and when autonomic versus conscious processing of interoceptive information occurs remains an intriguing area for further in-depth investigations [7].
Regulation of Interoception
The concept of regulating interoception via descending pathways has not been incorporated into most definitions of interoception, despite the well-established ability of the CNS to generate signals that regulate the internal state [41,42]. We will refer to these signals generated by the CNS to regulate the interoceptive processes, often in response to interoceptive input or cognitive and exteroceptive factors, as 'regulatory signals of interoception'. Neurons in the CNS involved in generating these regulatory signals can be called central regulators of interoception (Figure 3).
Figure 3 Illustrative Diagram of Sample Descending Neural Pathways of Interoception.
Regulatory signals of interoception, similar to interoceptive signals, can be transmitted via both non-neural (e.g., humoral) and neural pathways (e.g., cranial/vagal or spinal efferents) that target the peripheral organs [23,43]. In non-neural pathways, regulatory signals are delivered to the peripheral organ via the vascular or lymphatic systems and interact directly with the responding non-neural cells. In neural pathways, the final effectors, commonly called sympathetic or parasympathetic ganglion neurons, receive input from the brain through the spinal or vagal/cranial efferent nerves and directly synapse with internal organ non-neural cells [43] (Figure 3). It is worth noting that the non-neural and neural pathways may interact, such that a regulatory signal may be initially delivered through one pathway (e.g., a non-neural pathway) to impact on the other pathway (e.g., neural pathway) before reaching its final effector target.
The primary function of the regulatory signals of interoception is to regulate the generation and transmission of interoceptive signals of the targeted internal organs, which can be called effectors, and thus complete the circle of interoceptive processes. However, often the impact of the regulatory signals may be best measured by the responses or changes of function in the target organs. It is therefore sometimes impossible to distinguish regulatory interoceptive signals from body regulation. In our view, the inclusion of body regulation as a component of interoception science not only is necessary from a neuroanatomical perspective but also enables the development of more innovative methods to probe the functional impact of interoceptive processes, although we recognize that, in the current literature, many have described body regulation and interoception as separate terms and concepts.
Gaps and Challenges of Research in Interoceptive Neural Circuits
The picture of the interoceptive nervous system sketched in the previous text (Figures 1–3) is certainly incomplete. There are many gaps and methodological challenges in studying neural circuits of interoception at the neuroanatomical and functional levels.
The neuroanatomical techniques used to establish the knowledge base of interoception have primarily included histochemical and autoradiographic tract tracing, cytoarchitectonic analyses, and magnetic resonance imaging (MRI)-based diffusion tractography (DTI) [13,31,44,45]. Notably, tract-tracing studies show highly collateralized and interconnected networks across all nodes from the periphery to the cortex [22,46]. Additional experiments using tools such as transneuronal tracers and virus-based techniques will be necessary to establish a more detailed picture [17,22]. There are also multiple gaps between research focused on inputs from the periphery to the brainstem and those focused on the insular cortex [47–49]. For instance, more attention needs to be paid to examining the thalamic relay(s) [45]. In addition, much remains to be understood regarding the connections between the ascending and descending pathways, such as the hypothalamic endocrine connections and the connections from insula to other brain regions including somatosensory cortex and beyond. How do the neuronal and non-neural pathways connecting brain and periphery interact and influence each other [16]?
Converging and diverging projections are known to exist at all levels of interoception, but crucial cellular and synaptic-level information is missing. Experiments focused on the molecular-level specificity and diversity of neuronal types within the interoceptive system have only begun to emerge [17,50–52]. For instance, although attention has been paid to the different cytoarchitectonic regions of the insular cortex, is there a single representation of the body in the insula, or might there be multiple, overlapping maps [26,52]? Furthermore, it is important to note that the properties and cell types of the insular cortex appear to differ substantially across species, limiting the applicability of some findings from rodents to human interoceptive health [34]. At the periphery, transneuronal tracers have had limited application to date, consisting primarily of anterograde studies from scattered visceral organ systems [22,46,53–55]. Visceral afferents are low-density, thin, unmyelinated fibers that can be difficult to visualize [56]. Focusing on cell-type analysis and imaging of regions or nodes with a high density of neurons, such as various types of peripheral sensory ganglia, might offer an excellent opportunity to gain more comprehensive understanding [56–62]. Moreover, little is known about the diverse cell types involved in many of the key information-processing nodes for interoception from peripheral ganglia to cortex, whereas the complexity of circuits within these nodes, such as the brainstem and hypothalamus, remains largely unmapped.
Functional techniques have been mostly limited to early studies of evoked potentials in vagotomized animals and very few human lesion case reports. A more recent collection of resting state and task-based functional MRI (fMRI) studies in humans have revealed widespread viscera–brain coupling and placed insula function within the larger 'salience network' [27,30,33,36–38,63]. The insula and associated cortical and subcortical networks have been correlated with a wide range of possible functions related to interoception and allostasis, including visual perception, mental time-keeping, emotion, empathy, language and music perception, and self-awareness [23,26,27,34,38]. However, much remains to be understood about the many brain regions involved in interoception, especially whether specific neuronal populations in these regions function as interpreters, integrators, or regulators of interoceptive information.
Although interoception studies in humans currently rely primarily on fMRI blood oxygen level-dependent (BOLD) and DTI structural imaging [16] to provide crucial information about large-scale function, these methodologies are fundamentally correlative. In-depth, mechanistic studies that link anatomical findings to function will be important to substantiate the roles of brain regions and domains in interoception. In human subject research, non-invasive methodologies such as transcranial magnetic stimulation (TMS) and transcranial direct current stimulation (tDCS) may offer opportunities to assess the causal effects of brain regions in interoception [64]. However, the deep location of some of the brain regions involved in interoception, including the insular cortex, makes them difficult to image and stimulate [33]. Imaging studies in lesion patients, for instance AIC lesions, may also offer some causal evidence although they require caution in interpretation [24].
Functional Impact and Dynamics
Optimal sensing, interpretation, integration, and regulation of internal body signals, whether rising to the level of conscious awareness or not, are crucial for many essential physiological functions, such as breathing, eating, drinking, micturition, and maintaining body temperature, as well as for psychological experiences ranging from a variety of feelings and emotions to motivations and adaptive behaviors [3]. In addition, interoceptive and exteroceptive processes may interact to orchestrate complex physiological and behavioral functions, such as in the stress response [65]. The functions of interoception may range from essential bodily functions to high-level cognitive and emotional behaviors [66], and can be roughly grouped into two categories based on the direction of information flow.
The first category of interoceptive functions includes information flow from the body to the brain. For example, it is mostly unknown how affect – a brain state that can be described as a low-dimensional representation of high-dimensional behavioral state in the body [67] – is influenced by interoception. However, recent work on gut and hippocampal function [17], which tested the hypothesis that specific interoceptive signals are transported via the vagus nerve to the hippocampus to influence memory function, provides an elegant example on how one could begin to address this challenge.
The second category of interoceptive functions includes information flow from the brain to the body to exert bodily effects. The voluntary urination model is a good example. The target output of voluntary urination is the urethral sphincter, which is somatically controlled by the pontine micturition center (PMC, or Barrington's nucleus) of the brain [68], particularly corticotropin-releasing factor (CRF) labeled subpopulations of neurons in the PMC [69,70]. Stimulation of CRF-labeled neurons in the PMC caused immediate urination in mice [69], suggesting that the PMC neurons are sufficient to drive behavior. When neural activity was blocked in CRF-labeled neurons in the PMC, voluntary urination was inhibited. Often, this category of interoceptive function utilizes autonomic nervous system pathways, either sympathetic or parasympathetic.
In animal models, it is often possible to couple (i) neural activity manipulations, even at the single neuron level; (ii) neural activity measurements, such as electrophysiological or physiological assays, and/or in vivo neural imaging; with (iii) functional assessments to measure organ physiology and/or behavioral assays. Such experimental combinations have the potential to best support rigorous gain- and loss-of-function analyses of interoceptive neural circuits. By contrast, assessments of interoception in humans have been largely limited to a handful of approaches and remain mostly correlational. These approaches include heart-beat measures [71–73], skin conductance responses [74,75], and subjective self-report measures such as interoceptive accuracy or awareness [76]. More recently, microneurography has emerged as an objective tool to probe interoceptive responses [77,78]. If one includes body regulation as part of interoception research, additional physiological measures, such as blood pressure and baroreflex [79], may be explored to assess the functional impact of interoception. A key caveat in this context is that many organ functions are vital, making causal analyses challenging in human subject research of interoception. Furthermore, developing consistent experimental measures and metrics for comparable studies in humans and animal models will be crucial for translating insights across animal model and human studies.
Dynamic changes in interoceptive pathways over the lifespan remain largely unexplored. Several research groups have begun to study interoception in early life [80] and to examine how interoception changes with age [81–83], but relatively little is known about lifelong changes in interoception. One key challenge is the lack of a measure of interoceptive sensitivity that is suitable for non-verbal populations such as infants [80] or older adults with dementia [84]. In addition, conducting longitudinal studies and identifying milestones of interoception development will be necessary to uncover the changes in interoception across the lifespan.
Diseases and Disorders
Dysfunction of interoception has been increasingly recognized as an important component of many neurological, psychiatric, and behavioral disorders [3]. Altered structure, functional activity, or connectivity within the interoceptive neural network has been observed for instance in individuals with migraine and other types of chronic pain [85,86], alcohol and substance use disorders [87], anxiety, depression, and affective disorders [88,89], post-traumatic stress disorder [90], obsessive compulsive disorder [91], autism spectrum disorder (ASD) [92], eating disorders [93], somatic symptom disorders [94], stroke and neurodegenerative diseases [95,96], and may be a basis for comorbidity of neuropsychiatric and mental disorders [3,97].
For example, irritable bowel syndrome, characterized by visceral hypersensitivity, is associated with disruptions in the endocrine and immune systems in the gut as well as changes in the cortical neuronal network of sensorimotor, salience, emotion, and arousal [54] and disrupted modulation of insular cortex [98] in the human brain. Similarly, obesity is associated with disrupted interoceptive states from the gut to the brain such as altered functional connectivity of the insula [99] and distributed brain regions between the dorsal mid-insula, medial OFC, dorsal striatum, and ventral striatum [100].
Many psychiatric and neurodevelopmental disorders, such as schizophrenia [101], attention-deficit hyperactivity disorder [102], ASD, depression, and anxiety disorders, are not only linked to altered brain networks that are crucial for the integration of interoceptive signals for emotion processing and cognition but also exhibit a variety of physical symptoms [3]. For example, ASD is associated with dysregulated anterior insula connectivity and deficient emotional processing that may be due to aberrant prediction errors in interoceptive processing [72,103]. At the same time, individuals with ASD often show interoceptive changes such as increased sensitivity to pain and gastrointestinal symptoms [104].
Similarly, addiction to alcohol or other substances of abuse may cause adaptation in the interoceptive network which not only exacerbates stress and contributes to the alteration of emotion and reward processing but also gives rise to comorbid conditions through central and peripheral interactions [105,106]. A recent animal model study identified a genetic factor that links the neurocircuit adaptation of tobacco smoking to type 2 diabetes in humans [107], and provided evidence to show that the brain can control peripheral organ function while peripheral organs can regulate the addictive properties of nicotine. The gut–brain connection has also been shown to play a role in addictive behaviors and alcohol use disorders. For instance, ghrelin is a hunger hormone secreted in the stomach, and its concentration in individuals with alcohol use disorder correlates with alcohol drinking and predicts alcohol relapse [108,109]. In rodents, administration of ghrelin leads to an increase in alcohol reward and in alcohol intake, administration, and preference, whereas blockade of the ghrelin receptor resulted in reduction of these behaviors [108,109]. Moreover, evidence suggests that ghrelin, delivered through the vascular system, may mediate signaling from gut to brain via the vagus nerve [17,110].
A wide range of diseases and disorders are related to interoception [3,97], and pathophysiological and causal evidence has been generated for some of these conditions. However, it is important to note that, in most of them, the links to interoceptive dysregulation are largely symptomatic and descriptive. Future pathophysiological studies in clinical populations as well as in appropriate animal models are necessary for deepening the understanding of the possible outcomes of dysregulated interoceptive processing.
Manipulations and Interventions
The complex interplay between the ascending and descending pathways of the interoceptive system provides many potential routes and methods for targeted interventions in interoceptive dysfunction and related disorders. A multitude of approaches, ranging from non-invasive behavioral manipulations to specific pharmacological and neural stimulation interventions, have been considered for regulatory interoceptive processes [111].
In general, these approaches can be categorized into three groups – behavioral, neural stimulation, and pharmacological – each with advantages and limitations. Behavioral interventions, such as meditation and cognitive behavioral therapy (CBT), are the least invasive and generally the safest of the various interventions [112]. These approaches utilize exteroceptive routes (e.g., sound, vision, somatosensation, and cognitive influence) to trigger activity in the brain, and likely exert their effects on interoceptive body signals and function [113] through descending pathways. The limitations of these approaches include a lack of established therapeutic targets, relatively low potency, and delayed improvements [114]. Current neural stimulation approaches include TMS, tDCS, deep brain stimulation (DBS), vagal nerve stimulation (VNS), and transcutaneous electrical nerve stimulation (TENS) targeting peripheral nerves [115]. If the therapeutic targets are clear and specific, nerve stimulation can be a potent therapeutic with few side effects. However, therapeutic targets for most of these approaches have yet to be identified and validated. In addition, some approaches such as DBS and VNS may require invasive neurosurgery. Pharmacological interventions, such as blockade of the ghrelin receptor and/or of the peptide itself, represent promising approaches, although additional translational work is needed in that regard [108,109]. The pharmacological approach could be particularly effective for targeting interoceptors either within the brain, at the periphery, or along associated molecular signaling pathways. As in most pharmacological approaches, off-target effects and related side effects pose major challenges.
Regardless of the type of interventional approaches, identifying, assessing, and validating the therapeutic targets through rigorous mechanistic studies in both humans and appropriate animal models will be crucial for safety and efficacy in treating interoception-related diseases and disorders. In addition, group and individual variables including sex, gender, age, and social factors are important elements to consider for interventional studies.
Predictive Modeling
Computational modeling is an essential component of interoception research because it helps to formalize the empirical findings within a mathematical framework, as well as providing predictions for future experiments [116–118]. Several mathematical models are currently used to describe both the perceptual and body regulation aspects of interoception [117,119], although the sensory or perceptual aspects of interoception and the body regulation aspect are often modeled as separate processes. It may be desirable to develop computational models that could capture and describe the proposed framework of interoception in this article. In addition, modeling how interoceptive processes may be integrated with exteroceptive processes can provide a more comprehensive picture of how our nervous system interacts with the rest of the body to maintain function and support survival [119]. Modeling key parameters and outcomes that are accessible in experimental studies, such as 'interoceptive accuracy' and 'interoceptive awareness' [72,76], can greatly facilitate the development and validation of computational models and should be strongly encouraged.
Concluding Remarks
Although the foundations of the science of interoception were laid over 100 years ago, interoception research mostly regained momentum in recent years, partly because of the availability of high-resolution, multimodal tools for interrogating interoceptive processes. In this article we have outlined a proposed comprehensive framework of interoception science which may help to accelerate progress towards an integrative understanding of how we sense and regulate our internal states. Both conceptual and technical/methodological challenges remain (see Outstanding Questions), and our hope is that this review, together with other articles in this Special Issue, will offer stimulating ideas to enrich the emerging science of interoception.
Outstanding Questions
Interoceptors are molecular sensors or receptors in neurons that directly detect various interoceptive signals and transduce them into electrical, hormonal, or other non-neural signals to be integrated and interpreted by the brain. What are the molecular entities of these interoceptors? How can one systematically identify them?
What are the functional circuits of interoception? How do the central and peripheral nervous systems dynamically interact to support interoceptive processing in both physiological and pathological conditions?
What is the impact of central or peripheral disorders on interoceptive networks?
What are the effects of modulating interoceptive processes on diseases and disorders of the nervous system?
How can one develop more objective and quantitative assessments of interoception?
What are the most effective strategies for developing technologies and approaches to modulate interoceptive processes?
How do the interoceptive networks interact with other sensory, motor, reward, emotional, and cognitive/memory circuits to regulate nervous system diseases and disorders?
Disclaimer Statement
The content is solely the responsibility of the authors and does not represent the official views of the NIH or federal government.
Acknowledgements
The workshop entitled 'The Science of Interoception and Its Roles in Nervous System Disorders', that served as an important foundation for this review article, was supported by the NIH Blueprint for Neuroscience Research and funding contributions from NCCIH, NIAAA, NIDCR, and OBSSR of the NIH. We would like to thank Dr Paige Green from NCI, Dr Michael Oshinsky from NINDS, and Dr Marjorie Garvey from NIMH for their services in the workshop planning committee, the Directors and Coordination Committee members of the NIH Blueprint for Neuroscience Research Institute, Centers, and Offices for their support as well as thoughtful discussions on this effort, and Dr Emmeline Edwards from NCCIH for her support and feedback on the workshop as well as on the manuscript. In addition, we thank Catherine Law and Cindy Million for assistance with the revised manuscript. We would also like to thank all the invited speakers, panel discussants, and other workshop attendees for their intellectually stimulating presentations and discussions at the workshop. Lastly, we would like to thank Bryan Ewsichek from NCCIH for the graphic designs of Figures 1 and 3, and NIH Medical Arts for the graphic design of Figure 2 in this article.
References
Cameron, O.G.
Interoception: the inside story – a model for psychosomatic processes
Psychosom. Med. 2001; 63:697-710
Wiener, N.
The concept of homeostasis in medicine
Trans. Stud. Coll. Physicians Phila. 1953; 20:87-93
Khalsa, S.S. ...
Interoception and mental health: a roadmap
Biol. Psychiatry Cogn. Neurosci. Neuroimaging. 2018; 3:501-513
Tuthill, J.C. ∙ Azim, E.
Proprioception
Curr. Biol. 2018; 28:R194-R203
Shinder, M.E. ∙ Newlands, S.D.
Sensory convergence in the parieto-insular vestibular cortex
J. Neurophysiol. 2014; 111:2445-2464
Denton, D. ...
Neuroimaging of genesis and satiation of thirst and an interoceptor-driven theory of origins of primary consciousness
Proc. Natl. Acad. Sci. U. S. A. 1999; 96:5304-5309
Berntson, G.G. ∙ Khalsa, S.S.
Neural circuits of interoception
Trends Neurosci. 2021; 44:17-28
Ranade, S.S. ...
Piezo2 is the major transducer of mechanical forces for touch sensation in mice
Nature. 2014; 516:121-125
Chesler, A.T. ...
The role of PIEZO2 in human mechanosensation
N. Engl. J. Med. 2016; 375:1355-1364
Mei, N.
Recent studies on intestinal vagal afferent innervation. Functional implications
J. Auton. Nerv. Syst. 1983; 9:199-206
Janig, W.
Neurobiology of visceral afferent neurons: neuroanatomy, functions, organ regulations and sensations
Biol. Psychol. 1996; 42:29-51
Saper, C.B.
The central autonomic nervous system: conscious visceral perception and autonomic pattern generation
Annu. Rev. Neurosci. 2002; 25:433-469
Craig, A.D.
How do you feel? Interoception: the sense of the physiological condition of the body
Nat. Rev. Neurosci. 2002; 3:655-666
Barone, F.C. ∙ Zarco, I.
de Coronado, and M.J. Wayner, Gastric distension modulates hypothalamic neurons via a sympathetic afferent path through the mesencephalic periaqueductal gray
Brain Res. Bull. 1995; 38:239-251
Taher, J. ...
Central nervous system regulation of hepatic lipid and lipoprotein metabolism
Curr. Opin. Lipidol. 2017; 28:32-38
Critchley, H.D. ∙ Harrison, N.A.
Visceral influences on brain and behavior
Neuron. 2013; 77:624-638
Suarez, A.N. ...
Gut vagal sensory signaling regulates hippocampus function through multi-order pathways
Nat. Commun. 2018; 9:2181
Cechetto, D.F. ∙ Saper, C.B.
Evidence for a viscerotopic sensory representation in the cortex and thalamus in the rat
J. Comp. Neurol. 1987; 262:27-45
Cobos, I. ∙ Seeley, W.W.
Human von Economo neurons express transcription factors associated with layer V subcerebral projection neurons
Cereb. Cortex. 2015; 25:213-220
Champagne, D. ...
CRFergic innervation of the paraventricular nucleus of the rat hypothalamus: a tract-tracing study
J. Neuroendocrinol. 1998; 10:119-131
Iwai, H. ...
Ascending parabrachio-thalamo-striatal pathways: potential circuits for integration of gustatory and oral motor functions
Neuroscience. 2015; 294:1-13
Dum, R.P. ...
The spinothalamic system targets motor and sensory areas in the cerebral cortex of monkeys
J. Neurosci. 2009; 29:14223-14235
Gu, X. ...
Anterior insular cortex and emotional awareness
J. Comp. Neurol. 2013; 521:3371-3388
Wang, X. ...
Anterior insular cortex plays a critical role in interoceptive attention
Elife. 2019; 8, e42265
Penfield, W. ∙ Faulk, Jr., M.E.
The insula; further observations on its function
Brain. 1955; 78:445-470
Craig, A.D.
How do you feel – now? The anterior insula and human awareness
Nat. Rev. Neurosci. 2009; 10:59-70
Azzalini, D. ...
Visceral signals shape brain dynamics and cognition
Trends Cogn. Sci. 2019; 23:488-509
Kelly, C. ...
A convergent functional architecture of the insula emerges across imaging modalities
Neuroimage. 2012; 61:1129-1142
Gogolla, N.
The insular cortex
Curr. Biol. 2017; 27:R580-R586
King, A.B. ...
Human forebrain activation by visceral stimuli
J. Comp. Neurol. 1999; 413:572-582
Mufson, E.J. ∙ Mesulam, M.M.
Insula of the old world monkey. II. Afferent cortical input and comments on the claustrum
J. Comp. Neurol. 1982; 212:23-37
Khalsa, S.S. ...
The pathways of interoceptive awareness
Nat. Neurosci. 2009; 12:1494-1496
Uddin, L.Q. ...
Structure and function of the human insula
J. Clin. Neurophysiol. 2017; 34:300-306
Butti, C. ∙ Hof, P.R.
The insular cortex: a comparative perspective
Brain Struct. Funct. 2010; 214:477-493
Evrard, H.C. ...
Von Economo neurons in the anterior insula of the macaque monkey
Neuron. 2012; 74:482-489
Seeley, W.W. ...
Dissociable intrinsic connectivity networks for salience processing and executive control
J. Neurosci. 2007; 27:2349-2356
Kurth, F. ...
A link between the systems: functional differentiation and integration within the human insula revealed by meta-analysis
Brain Struct. Funct. 2010; 214:519-534
Kleckner, I.R. ...
Evidence for a large-scale brain system supporting allostasis and interoception in humans
Nat. Hum. Behav. 2017; 1, 0069
Mesulam, M.M. ∙ Mufson, E.J.
Insula of the old world monkey. III. Efferent cortical output and comments on function
J. Comp. Neurol. 1982; 212:38-52
Swanson, L.W. ∙ Petrovich, G.D.
What is the amygdala?
Trends Neurosci. 1998; 21:323-331
Li, M.M. ...
The paraventricular hypothalamus regulates satiety and prevents obesity via two genetically distinct circuits
Neuron. 2019; 102:653-667
Taggart, P. ...
Significance of neuro-cardiac control mechanisms governed by higher regions of the brain
Auton. Neurosci. 2016; 199:54-65
Janig, W. ∙ Habler, H.J.
Specificity in the organization of the autonomic nervous system: a basis for precise neural regulation of homeostatic and protective body functions
Prog. Brain Res. 2000; 122:351-367
Mesulam, M.M. ∙ Mufson, E.J.
Insula of the old world monkey. I. Architectonics in the insulo-orbito-temporal component of the paralimbic brain
J. Comp. Neurol. 1982; 212:1-22
Mufson, E.J. ∙ Mesulam, M.M.
Thalamic connections of the insula in the rhesus monkey and comments on the paralimbic connectivity of the medial pulvinar nucleus
J. Comp. Neurol. 1984; 227:109-120
Dum, R.P. ...
The mind–body problem: circuits that link the cerebral cortex to the adrenal medulla
Proc. Natl. Acad. Sci. U. S. A. 2019; 116:26321-26328
Ruggiero, D.A. ...
A role of insular cortex in cardiovascular function
J. Comp. Neurol. 1987; 257:189-207
Gehrlach, D.A. ...
Aversive state processing in the posterior insular cortex
Nat. Neurosci. 2019; 22:1424-1437
Livneh, Y. ...
Estimation of current and future physiological states in insular cortex
Neuron. 2020; 105:1094-1111
Kaelberer, M.M. ...
A gut–brain neural circuit for nutrient sensory transduction
Science. 2018; 361:26321-26328
Yang, L. ...
Transcriptomic landscape of von Economo neurons in human anterior cingulate cortex revealed by microdissected-cell RNA sequencing
Cereb. Cortex. 2019; 29:838-851
Evrard, H.C.
The organization of the primate insular cortex
Front. Neuroanat. 2019; 13:43
Rinaman, L. ∙ Schwartz, G.
Anterograde transneuronal viral tracing of central viscerosensory pathways in rats
J. Neurosci. 2004; 24:2782-2786
Rinaman, L.
Ascending projections from the caudal visceral nucleus of the solitary tract to brain regions involved in food intake and energy expenditure
Brain Res. 2010; 1350:18-34
McGovern, A.E. ...
Transneuronal tracing of airways-related sensory circuitry using herpes simplex virus 1, strain H129
Neuroscience. 2012; 207:148-166
Wang, W.
Optogenetic manipulation of ENS – the brain in the gut
Life Sci. 2018; 192:18-25
Udit, S. ∙ Gautron, L.
Molecular anatomy of the gut-brain axis revealed with transgenic technologies: implications in metabolic research
Front. Neurosci. 2013; 7:134
Kupari, J. ...
An atlas of vagal sensory neurons and their molecular specialization
Cell Rep. 2019; 27:2508-2523
Chang, R.B. ...
Vagal sensory neuron subtypes that differentially control breathing
Cell. 2015; 161:622-633
Williams, E.K. ...
Sensory neurons that detect stretch and nutrients in the digestive system
Cell. 2016; 166:209-221
Han, W. ...
A neural circuit for gut-induced reward
Cell. 2018; 175:665-678
e23
Rajendran, P.S. ...
Identification of peripheral neural circuits that regulate heart rate using optogenetic and viral vector strategies
Nat. Commun. 2019; 10:1944
Critchley, H.D. ...
Neural systems supporting interoceptive awareness
Nat. Neurosci. 2004; 7:189-195
Malik, S. ...
Deep TMS of the insula using the H-coil modulates dopamine release: a crossover [11C] PHNO-PET pilot trial in healthy humans
Brain Imaging Behav. 2018; 12:1306-1317
Mayer, E.A. ∙ Collins, S.M.
Evolving pathophysiologic models of functional gastrointestinal disorders
Gastroenterology. 2002; 122:2032-2048
Quigley, K.S. ...
Functions of interoception: from energy regulation to experience of the self
Trends Neurosci. 2021; 44:29-38
Barrett, L.F. ∙ Bliss-Moreau, E.
Affect as a psychological primitive
Adv. Exp. Soc. Psychol. 2009; 41:167-218
Pavcovich, L.A. ∙ Valentino, R.J.
Central regulation of micturition in the rat the corticotropin-releasing hormone from Barrington's nucleus
Neurosci. Lett. 1995; 196:185-188
Hou, X.H. ...
Central control circuit for context-dependent micturition
Cell. 2016; 167:73-86
Valentino, R.J. ...
Evidence for widespread afferents to Barrington's nucleus, a brainstem region rich in corticotropin-releasing hormone neurons
Neuroscience. 1994; 62:125-143
Brener, J. ∙ Ring, C.
Towards a psychophysics of interoceptive processes: the measurement of heartbeat detection
Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016; 371:20160015
Garfinkel, S.N. ...
Discrepancies between dimensions of interoception in autism: Implications for emotion and anxiety
Biol. Psychol. 2016; 114:117-126
Garfinkel, S.N. ∙ Critchley, H.D.
Threat and the body: how the heart supports fear processing
Trends Cogn. Sci. 2016; 20:34-46
D'Alonzo, M. ...
Modulation of body representation impacts on efferent autonomic activity
J. Cogn. Neurosci. 2020; 32:1104-1116
Zaman, J. ...
The use of stimulus perception to account for variability in skin conductance responses to interoceptive stimuli
Psychophysiology. 2020; 57, e13494
Mehling, W.
Differentiating attention styles and regulatory aspects of self-reported interoceptive sensibility
Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016; 371, 20160013
Park, J. ...
Abnormal sympathetic reactivity to the cold pressor test in overweight humans
Am. J. Hypertens. 2012; 25:1236-1241
Ackerley, R. ∙ Watkins, R.H.
Microneurography as a tool to study the function of individual C-fiber afferents in humans: responses from nociceptors, thermoreceptors, and mechanoreceptors
J. Neurophysiol. 2018; 120:2834-2846
Freeman, R. ∙ Chapleau, M.W.
Testing the autonomic nervous system
Handb. Clin. Neurol. 2013; 115:115-136
Maister, L. ...
Neurobehavioral evidence of interoceptive sensitivity in early infancy
Elife. 2017; 6, e25318
Khalsa, S.S. ...
Interoceptive awareness declines with age
Psychophysiology. 2009; 46:1130-1136
Mikkelsen, M.B. ...
Emotional reactivity and interoceptive sensitivity: exploring the role of age
Psychon. Bull. Rev. 2019; 26:1440-1448
Li, D. ...
Adolescent development of insula-dependent interoceptive regulation
Dev. Sci. 2017; 20, e12438
Toller, G. ...
Individual differences in socioemotional sensitivity are an index of salience network function
Cortex. 2018; 103:211-223
Maleki, N. ...
Female migraineurs show lack of insular thinning with age
Pain. 2015; 156:1232-1239
Suarez-Roca, H. ...
Contribution of baroreceptor function to pain perception and perioperative outcomes
Anesthesiology. 2019; 130:634-650
Migliorini, R. ...
What do you feel? Adolescent drug and alcohol users show altered brain response to pleasant interoceptive stimuli
Drug Alcohol Depend. 2013; 133:661-668
Avery, J.A. ...
Major depressive disorder is associated with abnormal interoceptive activity and functional connectivity in the insula
Biol. Psychiatry. 2014; 76:258-266
Grossi, D. ...
Altered functional connectivity of interoception in illness anxiety disorder
Cortex. 2017; 86:22-32
Simmons, A. ...
Initial evidence of a failure to activate right anterior insula during affective set shifting in posttraumatic stress disorder
Psychosom. Med. 2009; 71:373-377
Stern, E.R. ...
The buildup of an urge in obsessive-compulsive disorder: behavioral and neuroimaging correlates
Hum. Brain Mapp. 2020; 41:1611-1625
Hatfield, T.R. ...
Autism spectrum disorder and interoception: abnormalities in global integration?
Autism. 2019; 23:212-222
Donofry, S.D. ...
Alterations in emotion generation and regulation neurocircuitry in depression and eating disorders: a comparative review of structural and functional neuroimaging studies
Neurosci. Biobehav. Rev. 2016; 68:911-927
Harshaw, C.
Interoceptive dysfunction: toward an integrated framework for understanding somatic and affective disturbance in depression
Psychol. Bull. 2015; 141:311-363
Garcia-Cordero, I. ...
Feeling, learning from and being aware of inner states: interoceptive dimensions in neurodegeneration and stroke
Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016; 371, 20160006
Garcia-Cordero, I. ...
Stroke and neurodegeneration induce different connectivity aberrations in the insula
Stroke. 2015; 46:2673-2677
Bonaz, B. ...
Diseases, disorders, and comorbidities of interoception
Trends Neurosci. 2021; 44:39-51
Elsenbruch, S. ...
Patients with irritable bowel syndrome have altered emotional modulation of neural responses to visceral stimuli
Gastroenterology. 2010; 139:1310-1319
Kanoski, S.E. ∙ Grill, H.J.
Hippocampus contributions to food intake control: mnemonic, neuroanatomical, and endocrine mechanisms
Biol. Psychiatry. 2017; 81:748-756
Avery, J.A. ...
Obesity is associated with altered mid-insula functional connectivity to limbic regions underlying appetitive responses to foods
J. Psychopharmacol. 2017; 31:1475-1484
Ardizzi, M. ...
Interoception and positive symptoms in schizophrenia
Front. Hum. Neurosci. 2016; 10:379
Kutscheidt, K. ...
Interoceptive awareness in patients with attention-deficit/hyperactivity disorder (ADHD)
Atten. Defic. Hyperact. Disord. 2019; 11:395-401
Uddin, L.Q. ∙ Menon, V.
The anterior insula in autism: under-connected and under-examined
Neurosci. Biobehav. Rev. 2009; 33:1198-1203
Gu, X. ...
Heightened brain response to pain anticipation in high-functioning adults with autism spectrum disorder
Eur. J. Neurosci. 2018; 47:592-601
Herman, A.M. ∙ Duka, T.
Facets of impulsivity and alcohol use: what role do emotions play?
Neurosci. Biobehav. Rev. 2019; 106:202-216
Goldstein, R.Z. ...
The neurocircuitry of impaired insight in drug addiction
Trends Cogn. Sci. 2009; 13:372-380
Duncan, A. ...
Habenular TCF7L2 links nicotine addiction to diabetes
Nature. 2019; 574:372-377
Farokhnia, M. ...
Ghrelin: From a gut hormone to a potential therapeutic target for alcohol use disorder
Physiol. Behav. 2019; 204:49-57
Morris, L.S. ...
Stress, motivation, and the gut–brain axis: a focus on the ghrelin system and alcohol use disorder
Alcohol. Clin. Exp. Res. 2018;
Suarez, A.N. ...
Regulation of memory function by feeding-relevant biological systems: following the breadcrumbs to the hippocampus
Front. Mol. Neurosci. 2019; 12:101
Weng, H.Y. ...
Interventions and manipulations of interoception
Trends Neurosci. 2021; 44:52-62
Barrera, T.L. ...
A review of cognitive behavioral therapy for panic disorder in patients with chronic obstructive pulmonary disease: the rationale for interoceptive exposure
J. Clin. Psychol. Med. Settings. 2014; 21:144-154
Freeman, S.C. ...
Component network meta-analysis identifies the most effective components of psychological preparation for adults undergoing surgery under general anesthesia
J. Clin. Epidemiol. 2018; 98:105-116
Ehde, D.M. ...
Cognitive-behavioral therapy for individuals with chronic pain: efficacy, innovations, and directions for research
Am. Psychol. 2014; 69:153-166
Moisset, X. ...
Neurostimulation methods in the treatment of chronic pain
J. Neural Transm. (Vienna). 2020; 127:673-686
Seth, A.K. ...
An interoceptive predictive coding model of conscious presence
Front. Psychol. 2011; 2:395
Owens, A.P. ...
Interoceptive inference: from computational neuroscience to clinic
Neurosci. Biobehav. Rev. 2018; 90:174-183
Di Lernia, D. ...
Feel the time. Time perception as a function of interoceptive processing
Front Hum. Neurosci. 2018; 12:74
Petzschner, F.H. ...
Computational models of interoception and body regulation
Trends Neurosci. 2021; 44:63-76